Влечуги Съдържание Разпространение | Анатомия | Филогения и систематика | Произход и еволюция | Защитни механизми | Бележки | Литература | Вижте също | Външни препратки | НавигациявлечугиПрепоръка на Комисията от 18 юни 2007 година относно насоки за настаняване и грижи за животни, използвани за опитни и други научни целиLes vipères d'Europe de l'Ouest10.1016/j.gene.2006.09.026Lizards & Snakes Alive!10.1080/1063515060075539610.1111/j.1096-3642.1995.tb00932.xabstractarticleEarliest Evidence For ReptilesRainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica10.1130/G31182.1Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land10.1098/rsbl.2009.1024Complete mitochondrial genome suggests diapsid affinities of turtles10.1073/pnas.95.24.1422610.1038/384453a0PDF10.1098/rspb.1916.0012The Reptile Databasereptile (animal): BehaviourReptile and Amphibian Defense SystemsAnimal Planet: Ferocious CrocsPhylogeny and classification of amniotesAmniotaTree of Life web projectEMBL Reptile Database
Влечуги



 класгръбначни животничетирикракилюспипарафилетичнаАмниотаптицитебозайницитетоплообменоколната средамесояднияйцеснасящияйцеживораждащиживораждащикарбонземноводнидинозавриптерозавриихтиозавритеАнтарктидатропицитехабитатипустинигориблатаокеанигеконасоленоводния крокодилразредаптицитемонофилетичнаSauropsidaхерпетологиягръцкиАзияАфрикаЮжна Америкатропическите горипустинитенадморска височинаусойницатаЕвропаAmphisbaeniaевропейска блатна костенуркаанакондасмокообразнирекиезераМорските костенуркиокеаниМорските змииморетоГръбначният стълблатинскипрешленитеатласепистрофейеволюциятаглаватаребрагръдния кошГръдната косттазаопашкатаЧерепътземноводнитехрущялнебцелопаткатараменна костподбедрицаходилоноктиепидермисдермакожаталинеенебозайницитедерматаклеткипигментимеланиникаротиноидигуанинасветлинатахамелеонитенервната системаглавенгръбначен мозъкпродълговатия мозъкрибитеамфибиитенервасетивни органадишатбял дробустната кухиналаринксатрахеябронхиалвеолитемекочерупчестите костенуркиморските змиидиафрагмачерупкатамускулстуденокръвнисърдечно-съдовата системакръвообращениесърцетоартерииендокринната системадалакътХормонитеХипофизатаЛимфната системасинусихраносмилателната системаустаглъткахранопроводстомахтънкодебело червоклоакасляпо червоЧерният дробжлъчен мехурзадстомашна жлезаОтделителната системабъбрециуретрапикочен мехурсолиобмяната на веществатаполовата системаметаморфозакарбонапермаSynapsidaбозайнициАмниотаембрионалноразмножаванемезозоядинозавриптерозавриплезиозавримозазавриихтиозаврикредатерциерфилогениятатаксонмонофилияВкаменелоститерастителнояднихищнициSphenodontiaтриасНова ЗеландияЛюспеститеюрагущеризмиидвуходкиArchosauromorphaперммолекулярен часовникCrurotarsiалигаторовикрокодиловигавиаловилепидозавритеTree of Life Web ProjectMammaliaMesosauridaeMillerettidaeLanthanosuchidaeNyctiphruretiaPareiasauriaProcolophonoideaTestudinesCaptorhinidaeProtorothyrididaeAraeoscelidiaYounginiformesIchthyosauriaSauropterygiaLepidosauromorphaArchosauromorphaкарбонземноводниReptiliomorphaНова СкотиянасекомитеLabyrinthodontiaмасово измиранерастениеядствохищничествопермчерепносаочитегръбначния стълбкостенуркитебозайницичовеккарбончетирикракипеликозавридиметродонпермдинозавритекрокодилитезмиигущеритуатарипермЗемятаперм-триасово масово измиранетриасптерозавритемезозойската ераюраптициихтиозавритемезозойска еракредаМасово измиране Креда - Терциерморските костенуркилюспеститемезозойската ерабозайницикамуфлажкобритеотровницитеГеконитесцинковетеавтотомия
класгръбначни животничетирикракилюспипарафилетичнаАмниотаптицитебозайницитетоплообменоколната средамесояднияйцеснасящияйцеживораждащиживораждащикарбонземноводнидинозавриптерозавриихтиозавритеАнтарктидатропицитехабитатипустинигориблатаокеанигеконасоленоводния крокодилразредаптицитемонофилетичнаSauropsidaхерпетологиягръцкиАзияАфрикаЮжна Америкатропическите горипустинитенадморска височинаусойницатаЕвропаAmphisbaeniaевропейска блатна костенуркаанакондасмокообразнирекиезераМорските костенуркиокеаниМорските змииморетоГръбначният стълблатинскипрешленитеатласепистрофейеволюциятаглаватаребрагръдния кошГръдната косттазаопашкатаЧерепътземноводнитехрущялнебцелопаткатараменна костподбедрицаходилоноктиепидермисдермакожаталинеенебозайницитедерматаклеткипигментимеланиникаротиноидигуанинасветлинатахамелеонитенервната системаглавенгръбначен мозъкпродълговатия мозъкрибитеамфибиитенервасетивни органадишатбял дробустната кухиналаринксатрахеябронхиалвеолитемекочерупчестите костенуркиморските змиидиафрагмачерупкатамускулстуденокръвнисърдечно-съдовата системакръвообращениесърцетоартерииендокринната системадалакътХормонитеХипофизатаЛимфната системасинусихраносмилателната системаустаглъткахранопроводстомахтънкодебело червоклоакасляпо червоЧерният дробжлъчен мехурзадстомашна жлезаОтделителната системабъбрециуретрапикочен мехурсолиобмяната на веществатаполовата системаметаморфозакарбонапермаSynapsidaбозайнициАмниотаембрионалноразмножаванемезозоядинозавриптерозавриплезиозавримозазавриихтиозаврикредатерциерфилогениятатаксонмонофилияВкаменелоститерастителнояднихищнициSphenodontiaтриасНова ЗеландияЛюспеститеюрагущеризмиидвуходкиArchosauromorphaперммолекулярен часовникCrurotarsiалигаторовикрокодиловигавиаловилепидозавритеTree of Life Web ProjectMammaliaMesosauridaeMillerettidaeLanthanosuchidaeNyctiphruretiaPareiasauriaProcolophonoideaTestudinesCaptorhinidaeProtorothyrididaeAraeoscelidiaYounginiformesIchthyosauriaSauropterygiaLepidosauromorphaArchosauromorphaкарбонземноводниReptiliomorphaНова СкотиянасекомитеLabyrinthodontiaмасово измиранерастениеядствохищничествопермчерепносаочитегръбначния стълбкостенуркитебозайницичовеккарбончетирикракипеликозавридиметродонпермдинозавритекрокодилитезмиигущеритуатарипермЗемятаперм-триасово масово измиранетриасптерозавритемезозойската ераюраптициихтиозавритемезозойска еракредаМасово измиране Креда - Терциерморските костенуркилюспеститемезозойската ерабозайницикамуфлажкобритеотровницитеГеконитесцинковетеавтотомия
Влечуги
Направо към навигацията
Направо към търсенето
| Влечуги | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Класификация | ||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||
| Научно наименование | ||||||||||||||||||||||||||||
Laurenti, 1768[1] | ||||||||||||||||||||||||||||
| Разреди | ||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||
| Обхват на вкаменелости | ||||||||||||||||||||||||||||
вестфалий – настояще, 312–0 Ma ПреК К О С Д К П Т Ю К Пг Н | ||||||||||||||||||||||||||||
влечуги в Общомедия | ||||||||||||||||||||||||||||
Влечугите (Reptilia) са клас гръбначни животни, включват дишащи атмосферен въздух четирикраки, обикновено покрити с люспи. Макар и все още използвана в повечето съвременни системи за класификация, групата на влечугите е парафилетична, като обединява всички Амниота (гръбначни с амниотично яйце) с изключение на птиците, бозайниците и техни изчезнали роднини.[3]
Влечугите са животни с непостоянна телесна температура (пойкилотермни) – не произвеждат достатъчно топлина, за да поддържат постоянна телесна температура, а вместо това разчитат на топлообмен с околната среда, за да регулират вътрешната си температура, например чрез придвижване между сянка и слънце. Повечето влечуги са месоядни и яйцеснасящи, някои са яйцеживораждащи, а други са същински живораждащи.
Влечугите, възникват преди около 320 – 310 милиона години по време на периода на карбон. Еволюират от напреднали подобни на земноводни влечуги, които стават все по-адаптирани към живот на сушата. Много групи влечуги изчезват, включително динозаври, птерозаври и водните групи като ихтиозаврите.
Съвременните представители са разпространени на всички континенти, с изключение на Антарктида, като най-голямо разпространение имат в тропиците и субтропиците. Обитават подземни хабитати, пустини, гори, блата, океани и други. Размерите им варират от дължина 1,6 cm при гекона Sphaerodactylus ariasae до дължина 6 m и маса над 1000 kg при соленоводния крокодил (Crocodylus porosus).
Съвременните влечуги се разделят на 4 разреда:
Testudines (Костенурки) – около 300 вида;
Squamata (Люспести) – включва змии, гущери и други – около 7900 вида;
Rhynchocephalia (Клюноглави) – 2 вида;
Crocodylia (Крокодили) – 23 вида.
Въпреки че те имат люспи на краката си и снасят яйца, птиците са изключени от групата на влечугите, отчасти защото са топлокръвни. Затова не са включени в списъка по-горе. Въпреки това, тъй като някои влечуги са по-тясно свързана с птици, отколкото с други влечуги – крокодилите са по-тясно свързани с птици, отколкото с гущери. Някои учени ги групират в монофилетична група включваща и над 10 000 вида птици, известна като Sauropsida.
Науката изучаваща земноводни и влечуги се нарича херпетология (на гръцки: ἑρπετόν [херпетон] – „пълзящ“; и λογία [-логия] – „слово“, „наука“).
Съдържание
1 Разпространение
2 Анатомия
2.1 Скелет
2.1.1 Гръбначен стълб
2.1.2 Череп
2.1.3 Поясчета и крайници
2.2 Мускулатура
2.3 Кожа
2.4 Нервна система
2.5 Сетивни органи
2.6 Дихателна система
2.7 Кръвоносна система
2.8 Ендокринна система
2.9 Лимфна система
2.10 Храносмилателна система
2.11 Отделителна система
2.12 Полова система
3 Филогения и систематика
3.1 Съвременни групи
3.1.1 Произход на съвременните групи влечуги
3.2 Цялостна филогения
4 Произход и еволюция
4.1 Поява на влечугите
4.2 Възход на влечугите
4.3 Анапсиди, синапсиди и сауропсиди
4.4 Пермски влечуги
4.5 Мезозоят, ерата на влечугите
4.6 Изчезване на динозаврите
5 Защитни механизми
6 Бележки
7 Литература
8 Вижте също
9 Външни препратки
Разпространение |

Влечугите са се приспособили към множество среди, включително и океаните.
Влечугите са представени почти по цялото земно кълбо, с изключение на зоните, които е твърде студено – близо до полюсите. Тъй като те са студенокръвни животни, предпочитат по-скоро по-високи температури, а тяхното присъствие и разнообразие стават все по-голямо в близо до тропиците.[4] Така най-богатите континенти на влечуги са Азия, Африка и Южна Америка.
Влечугите са адаптирани към много различни местообитания. Техен дом са тропическите гори, където са отличават с много високо видово разнообразие, но също обитават и пустините, където има гущери и змии, които през деня се крият от горещините и излязат през нощта. В планинските райони гущерите се крият в купчини камъни, а някои змии са се специализирали в районите с висока надморска височина, като усойницата (Vipera ursinii), която се среща във високите планини на Европа при надморска височина до 2000 m.[5] Някои от влечугите се заравят и прекарват част от живота си под земята, както Amphisbaenia. Влечугите също са успели да колонизират водната среда: крокодилите, някои костенурки като европейска блатна костенурка и някои змии като анаконда, мокасинова водна змия (Agkistrodon piscivorus) и представители на смокообразни са обитатели на реки и езера. Морските костенурки са завладели всички океани по света и излизат на сушата само за да се размножават (да снесат яйцата си).[4]Морските змии проявяват по-високо ниво на адаптация, тъй като повечето от тях не се връщат на сушата и целият им жизнен цикъл преминава в морето. Много видове са се приспособили към живот по дърветата. Някои от тях могат да се движат от дърво на дърво като планират – летящи дракони (Draco Volans) и в по-малка степен някои летящи змии (Chrysopelea ornata).
Анатомия |

Скелет на Мегалания изчезнал от Австралия преди около 40 хил.години

Скелет на змия

Череп на крокодил
Скелет |
Гръбначен стълб |
Гръбначният стълб при влечугите е разделен на четири основни части: шийна (на латински: pars cervicalis), гръдно-поясна (pars thoracolumbalis), кръстна (pars sacralis) и опашна (pars caudalis).
Броят на прешлените варира при различните видове. Средният им брой е 50 – 80, но при змиите могат да достигнат до 140 – 435. Шийните прешлени са от 7 до 10 като първите два са атлас и епистрофей. Вторият се появява за първи път в еволюцията на гръбначните животни и прави възможно движението на главата наляво и надясно. При някои влечуги от последните три прешлена излизат чифтове шийни ребра. Гръдно-поясният отдел на гръбначния стълб е съставен от 16 до 25 прешлена като от всеки от тях излиза чифт ребра. Първите няколко чифта ребра заключват пространството на гръдния кош, който липсва при змиите. Гръдната кост е образувана от срастването на коремните части на няколко предни двойки ребра. Кръстната част на гръбначния стълб е представена само от два прешлена, които посредством издатъци се свързват с таза. Опашният отдел е съставен от няколко десетки постепенно намаляващи по размер прешлени, които към края на опашката съвсем изтъняват в миниатюрни костици.
При някои групи влечуги се наблюдават характерни отличия в устройството на гръбначния стълб. При змиите той видимо се дели само на две части – туловищна и опашна. При костенурките туловищната част на гръбначния стълб е сраснала с панцира и го прави неподвижен.
Череп |
Черепът при влечугите е вкостен в значително по-голяма степен в сравнение със земноводните. Единствено обонятелната капсула и слуховата област са изградени от хрущял. Слуховата област е съставена само от една кост наречена проотикум. Нова е и слъзната кост, която липсва при земноводните. За първи път при гущерите и крокодилите се появява твърдо небце. Лицевата и мозъчната част на черепа се залагат отделно в ембрионалното развитие, но по-късно се свързват в общо анатомично образувание. В състава на черепа освен първичните кости, които нормално го изграждат се включват и допълнителни кожни пластинки, които вторично вкостяват.
Поясчета и крайници |
Предното поясче е вкостено. Единствено при лопатката се наблюдава хрущялна супраскапула. Тазът е съставен от три чифтни кости свързани неподвижно помежду си. Крайниците са устроени подобно на тези при земноводните. Предните крайници са съставени от раменна кост, подмишница и длан с пръсти, а задните от бедро, подбедрица и ходило с пръсти. Пръстите накрая си имат нокти.
Мускулатура |
Мускулатурата в сравнение с тази на земноводните е по-добре развита, особено в главата, шията и крайниците. Представена е от дъвкателна, шийна, дихателна, а също така и флексори и екстензори осигуряващи процеса на движение на организма. Подкожната мускулатура позволява промяна на положението на роговите пластинки.
Кожа |

Линеене при Източна зелена мамба
Съставена е от епидермис и намираща се под него дерма. Епидермисът е най-външната покривка на кожата и често участва в образуването на различни кожни образувания. Такива са люспите при гущерите и змиите, костните пластини при крокодилите и панцира при костенурките. Смяната на епидермалния слой се осъществява при процеса на линеене. Отделя се частично или пълно от тялото на влечугото като отдолу остава нов слой, който бързо нараства и вкостява. Линеенето се осъществява до няколко пъти в годината. За разлика от бозайниците дермата е по-тънка. Наличието на епидермис позволява кожата на влечугите да издържа на сушата. Липсват и слизести жлези характерни за земноводните.
Във вътрешността на дермата се намират специални клетки наречени хроматофори. В тях се секретират пигменти: меланини и каротиноиди. Благодарение и на гуанина в тях те са способни и да отразяват светлината. Хроматофорите са способни да променят цвета на тялото за сравнително кратко време. Типичен пример за това е промяната на цвета на кожата при хамелеоните.
Нервна система |
Подобно на голяма част от гръбначните животни нервната система е представена от главен и гръбначен мозък. Главният мозък е разположен в черепната кутия. Отделните му части са почти еднакво развити. Характерна особеност е, че частите му не са разположени в права линия, а образуват лека чупка поради по-ниското разположение на продълговатия мозък. За разлика от рибите и амфибиите, при които главния мозък е от ихтиопсиден тип при влечугите той е от зауропсиден. Различават се пет части на главния мозък:
- Краен мозък – съставен е от две големи полукълба, разделени от дълбока мозъчна бразда. Повърхността на полукълбата е напълно гладка. При крокодилите се наблюдава струпване на мозъчни клетки, което се приема за зачатък на мозъчна кора.
- Междинен мозък – разположен е между предния и средния мозък. В горната му част се намира теменен орган, а в долната му част е хипофизата. Дъното на междинния мозък е заето от зрителните нерви и тяхната хиазма.
- Среден мозък – представен е от два големи предни хълма, представляващи зрителна част и два малки хълма в задната половина на мозъка. Зрителната кора е развита в значителна степен в сравнение с тази на амфибиите.
Малък мозък – покрива предната част на продълговатия мозък. Той е значително по-добре развит от този на земноводните.- Продълговат мозък – той прави характерна извивка във вентрална посока.
От главният мозък излизат 12 чифта черепно-мозъчни нерва.
Гръбначният мозък е удебелен в шийната и поясната област. От него излизат сегментални гръбначномозъчни нерви, които образуват раменни и тазови сплитове. Притежава и сиво мозъчно вещество под формата на буквата „Н“.
Сетивни органи |
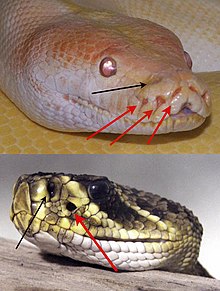
Терморецептори при различни видове змии

Око и слухов отвор при гущер
Влечугите притежават шест основни сетивни органа.
Очи. Устроени са по-сложно от тези на земноводните. В склерата се намира пръстен от твърди вкостени пластинки, от задната част на очната ябълка към склерата се вдава образувание наречено пектен, в ресничестото тяло се намира мускулатура, която позволява не само преместването на лещата, но и промяната на нейната форма. Очите са приспособени за възприемат образи във въздушна среда. Поради това са нужни и допълнителни спомагателни органи, които да запазват неговата цялост и функция. Такива са слъзните жлези, които посредством произвеждания от тях секрет не позволяват окото да изсъхне. Друг орган са клепачите. При някои змии се образува допълнителна ципа намираща се над окото. Поради характера на метаболизма си повечето влечуги са дневни животни. Ето защо е добре развито цветното зрение, което е най-чувствително към жълто-оранжевата част от спектъра на светлината.
Уши. Състоят се от вътрешно, средно и зачатък на външно ухо. Вътрешното ухо е съставено от утрикулус с три полуокръжни канала, сакулус с охлюв и мембрана базиларис. За разлика от зрението слухът няма първостепенна роля сред сетивата на влечугите. Те възприемат звуци в диапазона 20 – 6 000 Hz, но повечето от тях чуват добре само в диапазона 60 – 200 Hz (при крокодилите 100 – 3 000 Hz).- Обонятелен орган. Обонянието се извършва с помощта на хоаните и Якобсонов орган. За разлика от земноводните хоаните се намират близо до глътката и позволяват влечугото свободно да диша дори когато се храни.
- Орган на чувствителността към топлина (терморецептори). Намира се в лицевата ямка между очите и носа, симетрично от двете страни. Добре развит при змиите.
- Орган на вкуса. Вкусовите луковици са разположени основно в глътката.
- Осезателен орган. Осезателните образувания се намират под люспите и по езика.
Дихателна система |
Влечугите дишат с помощта на бял дроб като движенията на органа се осъществяват от налягането осъществено върху него от междуребрената и коремната мускулатура. В задната част на устната кухина е разположен отворът на ларинкса – глотис. В стените му има пръстеновиден и покривен хрущял. След ларинкса следва трахея, която в долната си част се разделя на бронхи за левия и десния дял на белия дроб. Подобно на земноводните белите дробове при влечугите имат торбовидна структура. Въпреки това обаче вътрешният им строеж е изключително сложен. Вътрешните стени на алвеолите са нагънати и прилепени една до друга и това увеличава значително дихателната повърхност.
Тъй като тялото е покрито с люспи, кожно дишане при влечугите липсва (с изключение на мекочерупчестите костенурки и морските змии) като белите дробове са единствения орган осъществяващ дишането. При крокодилите се е развила диафрагма, която допълнително помага при дишането. При костенурките дишането е ограничено от черупката. Установено е, че белият дроб е обвит от тънък мускул, който при съкращаването си помага при дишането.
Кръвоносна система |
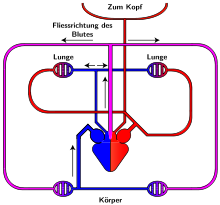
Схема на кръвообращението при влечугите
Подобно на рибите и земноводните, влечугите също са студенокръвни животни. Това се дължи в най-голяма степен от устройството на сърдечно-съдовата система и типа на кръвообращение.
Отвън сърцето е обвито с перикард. Съставено е от една камера и две предсърдия. Чрез непълна преграда камерата е разделена на лява и дясна половина. При крокодилите има почти пълна преграда, но с малък отвор. Между предсърдието и камерата се намира по едно полулунна клапа. От камерата излизат три самостоятелни артерии:
- Артерията излизаща от лявата част на камерата носи окислена кръв и дава разклонения за лява и дясна сънна артерия и продължава към дясната дъга на аортата.
- Артерията излизаща от дясната част на камерата носи смесена кръв и образува лявата дъга на аортата.
- Третата артерия излиза също от дясната половина на камерата и носи венозна кръв. Разделя се на дясна и лява белодробна артерия.
След като се отдели окислената кръв към главата двата дяла на аортата се съединяват в една нечифтна гръбна аорта и продължава назад като дава различни разклонения към органите на тялото. Така смесената и артериалната кръв допълнително се смесват в общата аорта.
Ендокринна система |
От ендокринната система далакът е добре развит. Хормоните на тиреоидната жлеза контролират отделянето на роговата обвивка. Хипофизата е също добре развита.
Лимфна система |
Лимфната система се състои от подкожни синуси и лимфни сърца в основата на опашката.
Влечугите имат сложно изградена лимфна система, чийто многобройни съдове обгръщат вените. Под гръбначния стълб преминава главният лимфен съд, който в горната си част се разклонява, а двата му клона се вливат в горните кухи вени. В задната част на тялото се намират други големи съдове както и лимфните сърца, които изпомпват лимфата в тазовите вени.[6]
Храносмилателна система |
Благодарение на разнообразието от хранителни ресурси, с които влечугите се хранят се появява и по-ясна диференциация на отделите от храносмилателната система. Съставена е от уста, глътка, хранопровод, стомах, тънко, дебело черво и клоака. Между тънкото и дебелото черно се намира сляпо черво, което липсва при земноводните. Черният дроб е с жлъчен мехур, притежават и задстомашна жлеза.
Отделителна система |
Отделителната система при влечугите е представена от бъбреци, уретра и пикочен мехур.
За разлика от рибите и амфибиите, при които бъбреците трябва да се справят с отделянето на голямо количество вода от организма, при влечугите вследствие на преминаването към сухоземен начин на живот бъбреците трябва да филтрират кръвта като отделят голямо количество соли и непотребни вещества, образувани в резултат на обмяната на веществата. При първите бъбреците са от типа мезонефрос. При влечугите бъбреците вече са от типа заден бъбрек (метанефрос, meta – „след“, nephros – „бъбрек“). От мезонефроса еволюционно се запазват само каналите, които остават като части на половата система. При метанефроса се оформят малпигиеви телца. Те нямат каналчета, които започват от тях (нефростоми) и са разположени близо едно до друго. Поради това бъбрекът е къс и компактен.
Като изходен канал служи отклонение от Волфовия канал, наречено уретра. Тя е тънкостенна и се отваря на вентралната страна на клоаката. При някои влечуги (крокодили, змии, някои гущери) уретрата е слабо развита.
Полова система |

Чифтосващи се гущери от вида Зелен анолис (Anolis carolinensis)
Влечугите са разделнополови организми.
Женска полова система. Състои се от яйчници, които нямат връзка с яйцепроводите. Оплождането се извършва в яйцепроводите. Последните се отварят в клоаката.
Мъжка полова система. Съставена е от семенници с каналчета, които излизат от тях и семепровод, който се отваря в уретрата преди клоаката. При гущерите в клоаката се намират два копулационни израстъка, по които се стичат сперматозоидите. Те нямат структура на пенис, макар че служат при копулация. За пръв път пенис се появява при крокодилите.
Влечугите снасят яйца. Някои са яйцеживородни или живородни. Развитието им е без метаморфоза.
Филогения и систематика |
Първите влечуги се появяват преди около 300 млн. години в края на карбона и началото на перма. Заедно със Synapsida (включващи появилите се по-късно бозайници), те произлизат от Амниота, четирикраките гръбначни с амниотично яйце, позволяващо сухоземно ембрионално развитие. За разлика от земноводните, влечугите са напълно приспособени към живота на сушата. С ефикасни бъбреци, намаляващи загубата на вода, непромокаема кожа и възможност за независимо от водата размножаване, те завладяват сушата, въздуха и нахлуват обратно към водата, постигайки най-голямото си разнообразие в „ерата на влечугите“, мезозоя. Повечето от тогавашните представители (динозаври, птерозаври, плезиозаври, мозазаври, ихтиозаври и др.) измират най-късно на границата креда/терциер (преди 65 млн. години).
През последните години филогенията на влечугите е обект на редица спорове. Много от старите наименования се избягват или получават нови значения. Разглеждани като монофилетичен таксон, самите влечуги получават названието Sauropsida, включващо (последния общ предшественик и потомците му, включително потомците на днешните) „влечуги и всички други амниоти, по-близки с тях, отколкото с бозайниците“ (Gauthier 1994). От друга страна понятието Reptilia получава и друго значение, като наименование на таксона, включващ последния общ предшественик на запазените видове влечуги (Gauthier et al., 1988).
Съвременни групи |
Следният списък опростено представя връзките между съвременните групи Sauropsida, според най-разпространената филогенетична систематика:
Sauropsida- Reptilia (Linneaus sensu Gauthier, 1984)
- Anapsida („без апсида“, без черепен отвор зад окото)
Testudines (=Testudinata=Chelonia) (Костенурки)
- Romeriida
- Diapsida („две апсиди“, два черепни отвора зад окото)
- Lepidosauromorpha
Lepidosauria (Лепидозаври)
Squamata (Люспести)
Rhynchocephalia (вкл. туатара)
- Archosauromorpha
Archosauria (Архозаври)- Crocodylomorpha
Crocodylia (Крокодили)
- Dinosauria
Aves (Птици)
- Crocodylomorpha
- Lepidosauromorpha
- Diapsida („две апсиди“, два черепни отвора зад окото)
- Anapsida („без апсида“, без черепен отвор зад окото)
- Reptilia (Linneaus sensu Gauthier, 1984)
Противно на някои разпространени твърдения, произходът на птиците от влечугите не се оспорва от днешната наука. Единствено стриктното настояване за монофилия на таксоните, а от там и необичайното класифициране е сравнително ново явление.
Произход на съвременните групи влечуги |
- Костенурки
Костенурките, (Testudines) може да са се развили от анапсиди, но точният им произход е неизвестен и силно обсъждан. Вкаменелостите датират от около 220 милиона години назад и споделят забележително сходни характеристики. Тези първи костенурки притежават същото устройство на тялото като всички съвременни костенурки и са предимно растителноядни. Срещат се и малко видове хранещи се единствено с малките морски организми. Смята се, че запазената им марка, черупката, е еволюирала от разширения на гръбнака, които срастват с разширения на ребрата. Това се подкрепя от фосили на Odontochelys semitestacea, който има непълна черупка, която произхожда от ребрата и гръбнака. Този вид също така има зъби в клюна си, което дава подкрепа на хипотезата, че е преходно изкопаемо, въпреки че това твърдение все още е спорно. Тази обвивка се развива, за да се предпази костенурката от хищници, но също така прави тромави сухоземните видове и ги забавя при движение. Това е причината много видове да изчезнат в последно време. Причина за това са неавтохтони видове, привнесени от човека, конкуриращи се с бавните костенурки за храна и липсата на възможност да избягат от хората. Това води до наличието на много застрашени видове в този разред.
- Клюноглави
Sphenodontia се появяват в средата на триас, но до наши дни е оцелял само от един род Sphenodon, който включва само два застрашените вида от Нова Зеландия и някои други по-малки околни острови. Тяхната еволюционна история е изпълнена с много видове.
- Люспести
Най-новият разред на влечугите. Люспестите се отличават по подвижната квадратна кост (като им позволява движение на горната челюст) и наличието на рогови люспи. Те произхождат от началото на юра и са съставени от три подразреда: гущери (Lacertilia), змии (Serpentes) и двуходки (Amphisbaenia). Въпреки че те са най-младия разред, Squamata съдържат повече видове, отколкото всеки от другите разреди влечуги. Те се монофилетична група, която е сестрина група (клон) на туатарите (Sphenodontia). От своя страна люспестите и туатарите заедно са сестрина група на крокодили и птици, или това са съвременните представители на архозаврите (Archosauromorpha). Въпреки че вкаменелости от люспести се появяват в началото на юра, митохондриални филогенетични изследвания предполагат, че те са еволюирали в края на перм. Повечето еволюционни взаимоотношения в рамките на разреда не са напълно изяснени, като връзката на змиите с другите групи е най-проблемна. От морфологично данни се смята, че игуановите гущери са се отделили от другите люспести много рано, но последните резултати от молекулярни филогенетични проучвания, както на митохондриите така и на ядрената ДНК, не са в подкрепа на това ранно разделяне.[7] Тъй като змиите имат по-бърз молекулярен часовник от други люспести[7] и има малко фосили от ранни змии и прародители на змиите,[8] е трудно да се установят връзките между змии и другите групи люспести.
- Крокодили
Първите организми, които показват сходни характеристики с крокодилите са Crurotarsi, които се появяват през началото на триас, преди 250 млн. години. Това бързо води развитието на клона на същинските крокодили (Eusuchia) преди 220 млн. години, което в крайна сметка дават началото на разреда крокодили, първият от които е възникнал преди около 85 милиона години по време на късната креда. Най-ранните изкопаеми доказателства за eusuchia е на вида Isisfordia. Ранните видове, основно се хранят с риба и растителност. Те са били сухоземни, повечето са с дълги крака (при сравнение със съвременните крокодили) и много от тях бяха двуноги. С увеличаване на разнообразието се появяват много върховни хищници, всички от които сега са изчезнали. Съвременните крокодили еволюират чрез появата на специфични еволюционни черти. Пълна загуба на двуногите, за сметка на като цяло ниската позиция на четириногите за лесно и по-малко забележимо влизане на телата във вода. Формата на черепа/челюстта се променят, за да се даде възможност допълнително да се хванат заедно със сочещите напред ноздри и очи. Развиват мимикрия необходима им за хищническото поведение на дебнене. Гърба на всички крокодили прилича на някаква плаваща трупа, заедно с общата окраска в кафяво и зелено за да имитират мъх или дърво. Опашката им има форма на гребло, способстваща за увеличаване на скоростта на плуване. Единствените оцелели групи в този разред са алигаторови, крокодилови и гавиалови.
Цялостна филогения |
Освен костенурките, всички днешни влечуги, както и птиците, спадат към Diapsida. За цялостен анализ на групата се налага и разглеждането на изчезналите видове.
До преди десетина години преобладава разделянето на влечугите по следния начин:
- Sauropsida
- Reptilia (по горната дефиниция: включващ последния общ предшественик на днешните влечуги и птици), обединяващ:
- Anapsida (Captorhinidae † и Testudines)
- Romeriida (Diapsida и парафилетичният Protorothyrididae †)
- „паравлечуги“, вкл. Millerettidae †, Pareiasauria †, Procolophonoidae †, Mesosauridae †
- Reptilia (по горната дефиниция: включващ последния общ предшественик на днешните влечуги и птици), обединяващ:
Така Testudines (костенурките) са съседни на Captorhinidae анапсиди.
Съществува обаче и хипотеза, че костенурките не са същински анапсиди, а са загубили черепните си отвори след развитието си от диапсиди. Ако е вярна, костенурките най-вероятно са много по-близки до лепидозаврите, отколкото до други днешни групи.
Друга тенденция е утвърждаването на „паравлечугите“ като монофилетичен таксон Parareptilia, който да включва и костенурките. Скорошни изследвания показват, че „паравлечугите“ е много вероятно да са изцяло анапсиди, а Captorhinidae да е по-правилно да се причислят към Romeriida, вместо като най-близки на костенурките. Това води до следната систематика:
- Reptilia (по дефиниция включващ последния общ предшественик на днешните влечуги и птици)
- Anapsida (с костенурките и „паравлечугите“)
- Romeriida (Diapsida, Captorhinidae † и парафилетичният Protorothyrididae †)
Това би отъждествило Reptilia (по горната дефиниция) със Sauropsida (по дефиницията от по-рано), тъй като последният общ предшественик на днешните влечуги в този случай е и последният общ предшественик на всички влечуги (Sauropsida).
Кладограма илюстрираща родословното дърво на влечуги. Следва опростена версия на отношенията, открити от Laurin and Gauthier (1996), представени като част от проекта Tree of Life Web Project,[9] с информация за връзките на най-примитивните влечуги според Мюлер и Reisz (2006).[10][Забележка 1]
| Amniota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
↑ Показаната филогения не отразява съвременните молекулярни доказателства причисляващи морските костенурки към Diapsida.
Произход и еволюция |
Поява на влечугите |
Първите влечуги се появяват в края на карбон, преди около 320 – 310 милиона години и произлизат от древните земноводни от групата Reptiliomorpha.[11] Най-старите известни следи от влечуги са отпечатъци от стъпки във фосилни пластове в Нова Скотия на възраст 315 милиона години,[12] за които се смята, че са оставени от представител на рода Hylonomus.[13]
Това е най-старото известно влечуго, подобно е на гущер с дължина 20 – 30 cm и има многобройни остри зъби, които показват, че основната им храна са насекомите.[14] Други ранни влечуги, които силно наподобяват по устройство и вероятен начин на живот Hylonomus са Westlothiana и Paleothyris. До малкият ледников период в края на карбона влечугите остават малка част от фауната, засенчвани от значително по-едрите земноводни Labyrinthodontia.
Възход на влечугите |
Края на карбон е съпроводен с рязко засушаване на климата (Карбонски колапс на влажните гори), водещо след себе си масово измиране на сухоземни организми.[15] Тази внезапна промяна засяга няколко големи групи. Земноводните са частично унищожени, но влечугите се справят по-добре, тъй като са екологично по-приспособени към сухите условия, които настават. За да съществуват като вид земноводните трябва да имат достъп до вода, където да снасят яйца. За разлика от тях влечугите, чиито яйца притежават черупка, която им позволява да бъдат снасяни на сушата, са по-добре адаптирани към новите условия. Влечугите завладяват нови ниши с по-бързи темпове, отколкото преди засушаването, при това много по-бързо от земноводните. Попаднали при нови условия бързо успяват да развият адаптивни приспособления и нови стратегии за хранене – растениеядство и хищничество. Преди това влечугите са били насекомоядни и рибоядни.[15] От този момент нататък, влечугите доминират съобществата и имат по-голямо разнообразие от земноводните, подготвяйки сцената за мезозойската „Ерата на динозаврите“.[16] Едно от най-известните ранните влечуги е Mesosaurus от началото на перм, който се е завърнал към вода и се храни с рибата.
Анапсиди, синапсиди и сауропсиди |

A=Анапсиди (Anapsida), B=Снапсиди (Synapsida), C=Диапсиди (Diapsida)
Първите влечуги са били анапсидни, притежавали масивен череп, без отвори за носа, очите или гръбначния стълб [17]. Счита се, че костенурките са единствените оцелели анапсидни влечуги, тъй като техният череп е изграден на този принцип. Съществува обаче и алтернативна теория, според която костенурките всъщност са еволюирали към това по-примитивно състояние, като приспособление за укрепване на бронята си [11]. И двете теории имат солидни доказателства, поради което няма единно мнение [18][19][20].
Скоро след появата на първите влечуги, еволюцията им се разделя на два отделни клона [21]. Единият клон представляват т.нар. синапсиди (включва всички изчезнали висши влечуги, както и съвременните бозайници, в това число и човек), които притежават отвори в черепния покрив, точно зад всяко око. Вторият клон – динапсидите притежава освен отвор зад всяко око още един разположен над първия. Функцията на тези отвори е олекотяване на черепа и приспособяването на челюстните мускули към по-здрава захапка [17].
Диапсидите и по-късните анапсиди се обединяват в т.нар. „същински влечуги“ или сауропсиди [22].
Пермски влечуги |
В края на карбон, влечугите представляват господстващите четирикраки животни. По времето когато сухоземните рептилоподобни лабиринтодонти все още съществуват, се появяват и същински сухоземни гиганти под формата на пеликозаври, като например едафозавъра (Edaphosaurus) и хищният диметродон (Dimetrodon). През среден перм, климатът става значително по-сух, което е причина за промяна на фауната – примитивните пеликозаври се изместват от по-съвършените терапсиди [23].
Анапсидните влечуги, чиито масивен череп няма посторбитални отвори, продължават съществуването си и дори преживяват разцвет по време на целия перм. Пареязаврите достигат гигантски размери през късния перм, но изчезват в края на периода с единствено изключение в съвременните костенурки [23].
По-рано през перм, от диапсидните влечуги се обособяват две основни направления – архозаври (предшествениците на динозаврите и крокодилите) и лепидозаврите (предшествениците на съвременните змии, гущери и туатари). Двете групи остават гущероподобни и с относително малки размери по време на целия перм.
Мезозоят, ерата на влечугите |
В края на перм настъпва едно от най-големите измирания на живи същества в историята на Земята – т.нар. перм-триасово масово измиране. Повечето от ранните анапсиди и синапсиди отмират и биват изместени от архозавроподобните диапсиди. Архозаврите се характеризират с дълги задни крайници и изправена поза, което им придава вида на дългокраки крокодили. Архозаврите стават господстваща група по време на триас и дават началото на повечето добре познати динозаври, птерозаврите, крокодилите и фитозаврите. Някои от динозаврите се превръщат в едни от най-големите сухоземни животни, живели някога на планетата, което е причина мезозойската ера да се нарича „ерата на динозаврите“. Някои динозаври дават началото на малки форми, включително и на пернати „тероподи“. През средата на юра се появяват и първите птици [23].
Диапсидите дават началото на водните влечуги [24]. Водните влечуги от начало са примитивни, като ранните сауроптериги през ранния триас и ихтиозаврите през следния триас. Мезозаврите се появяват през мезозойската ера през креда.
Терапсидите са под еволюционен натиск през ранния мезозой и се развиват като малки и нощни форми, като по-късно през юра те дават началото на първите бозайници.
Като цяло може да се каже, че влечугите са в разцвет през цялата мезозойска ера.
Изчезване на динозаврите |
В края на креда настъпва широкомащабно измиране на живи същества, известно като Масово измиране Креда - Терциер. От огромните морски влечуги оцеляват единствено морските костенурки, а от динозаврите оцеляват някои малки пернати тероподи като птиците. Краят на „ерата на динозаврите“ води до началото на „ерата на бозайниците“. Въпреки смяната на доминиращите форми, еволюцията на влечугите продължава и през ценозоя, като например люспестите преживяват най-голямото си развитие и видообразуване – по-голямо от това през мезозойската ера. Днес люспестите представляват около 90% от съвременните влечуги [25]. Влечугите наброяват около 8700 съвременни вида [25], което може да се сравни с около 5400-те вида бозайници.
Защитни механизми |

Гекон от вида Phelsuma deubia на фона на палмово листо
Много дребни влечуги, живеещи на земята или във водата, като например змиите и гущерите са уязвими от различни видове хищници. Най-честата форма на защита обаче си остава бягството и избягването на конфронтация [26]. При първите признаци на опасност, повечето змии и гущери се скриват под земята, а други влечуги като водните костенурки и крокодилите се гмуркат надълбоко. Влечугите могат да избягват нападатели чрез камуфлаж – с използването на шарки на сивото, зеленото и кафявото, влечугите се прикриват чудесно сред околната среда на естествените им местообитания [27].
Ако опасността е изненадваща или неприятелят е настоятелен, тогава някои влечуги като крокодилите, костенурките, някои гущери и змиите съскат силно с надеждата да отблъснат врага. Гърмящите змии тръскат върха на опашката си, при което се чуват серии от тракания. Ако всичко това не помага то различните видове използват допълнителни защитни стратегии.
Змиите притежават сложно поведение, когато са атакувани. Някои видове се изправят и надуват кожата около врата си в опит да се представят за по-големи и плашещи. Неуспехът на тази тактика, принуждава кобрите, отровниците и сходните видове да използват змийската си отрова. Отровата е видоизменена слюнка, която са инжектира в жертвата посредством отровни зъби.

Гекон с откъсната опашка
Когато крокодилите са притеснени за безопасността си, те показват зъбите си и жълтия си език. Ако това не помага, то крокодила е видимо притеснен и започва да съска. При неуспех и на тази тактика, крокодилите започват да сменят позата си многократно и бързо, като същевременно издуват тялото си за постигане на по-големи размери. Последната стъпка е атака на неприятеля.
Някои видове влечуги опитват да хапят, други използват главите си като чукове за буквално смазване на опонента си, трети атакуват от разстояние със засилка и сблъсък [28].
Геконите, сцинковете и други гущери използват необикновената тактика на пречупване на опашката (т.нар. автотомия), ако биват хванати или дори при опасност да бъдат хванати. Откъснатата опашка продължава да мърда, което обърква неприятеля за местоположението на гущера и позволява на жертвата да избяга. Тези животни могат да регенерират частично опашката си за няколко седмици. Новите сегменти съдържат хрущяли за сметка на костите от гръбначния стълб, а кожата на новата опашка може да е с различна шарка от останалата част на тялото.
Бележки |
↑ указано в: Обединена система за таксономична информация, дата на публикуване: 2000, дата на проверка: 19 септември 2013
↑ Препоръка на Комисията от 18 юни 2007 година относно насоки за настаняване и грижи за животни, използвани за опитни и други научни цели
↑ Tudge, Colin. The Variety of Life. Oxford University Press, 2000. ISBN 0198604262.
↑ аб Mark O'Shea et Timothy R. Halliday. Reptiles et amphibiens. 2005. ISBN 2035604230. O'Shea2005. с. 256.
↑ Dieter Zorn. Les vipères d'Europe de l'Ouest. // Посетен на 21 septembre 2011.
↑ Пешев 2002.
↑ аб Kumazawa, Yoshinori. Mitochondrial genomes from major lizard families suggest their phylogenetic relationships and ancient radiations. // Gene 388 (1 – 2). 2007. DOI:10.1016/j.gene.2006.09.026. с. 19 – 26.
↑ Lizards & Snakes Alive!. // American Museum of Natural History. Посетен на 25 декември 2007.
↑ Laurin, M. and Gauthier, J.A. (1996). „Amniota. Mammals, reptiles (turtles, lizards, Sphenodon, crocodiles, birds) and their extinct relatives.“ Version 1 January 1996. http://tolweb.org/Amniota/14990/1996.01.01 in The Tree of Life Web Project, http://tolweb.org/
↑ Muller, J. and Reisz, R.R. (2006). „The phylogeny of early eureptiles: Comparing parsimony and Bayesian approaches in the investigation of a basal fossil clade.“ Systematic Biology, 55(3):503 – 511. doi:10.1080/10635150600755396
↑ аб Laurin, M. и др. A reevaluation of early amniote phylogeny. // Zoological Journal of the Linnean Society 113. 1995. DOI:10.1111/j.1096-3642.1995.tb00932.x. с. 165 – 223. (abstract)
↑ Falcon-Lang, H.J., Benton, M.J. & Stimson, M. (2007): Ecology of early reptiles inferred from Lower Pennsylvanian trackways. Journal of the Geological Society, London, 164; no. 6; pp 1113 – 1118. article
↑ Earliest Evidence For Reptiles
↑ The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals. London, Marshall Editions, 1999. ISBN 1-84028-152-9. с. 62.
↑ аб Sahney, S., Benton, M.J. & Falcon-Lang, H.J.. Rainforest collapse triggered Pennsylvanian tetrapod diversification in Euramerica. // Geology 38 (12). 2010. DOI:10.1130/G31182.1. с. 1079 – 1082.
↑ Sahney, S., Benton, M.J. and Ferry, P.A.. Links between global taxonomic diversity, ecological diversity and the expansion of vertebrates on land. // Biology Letters 6 (4). 2010. DOI:10.1098/rsbl.2009.1024. с. 544 – 547.
↑ аб Alfred Romer|Romer, A.S. & T.S. Parsons. 1977. The Vertebrate Body. 5th ed. Saunders, Philadelphia. (6th ed. 1985)
↑ Benton, M. J.. Vertebrate Paleontology. 2nd. London, Blackwell Science Ltd, 2000. ISBN 0632056142., 3rd ed. 2004 ISBN 0-632-05637-1
↑ Zardoya, R. и др. Complete mitochondrial genome suggests diapsid affinities of turtles. // Proc Natl Acad Sci U S A 95 (24). 1998. DOI:10.1073/pnas.95.24.14226. с. 14226 – 14231.
↑ Rieppel, O. и др. Turtles as diapsid reptiles. // Nature 384. 1996. DOI:10.1038/384453a0. с. 453 – 455.
↑ van Tuninen, M. & Hadly, E.A. (2004): Error in Estimation of Rate and Time Inferred from the Early Amniote Fossil Record and Avian Molecular Clocks. Journal of Mulecular Biology, no 59: pp 267 – 276 PDF
↑ Goodrich, E.S.. On the classification of the Reptilia. // Proceedings of the Royal Society of London 89B. 1916. DOI:10.1098/rspb.1916.0012. с. 261 – 276.
↑ абв Colbert, E.H. & Morales, M. (2001): Colbert's Evolution of the Vertebrates: A History of the Backboned Animals Through Time. 4th edition. John Wiley & Sons, Inc, New York – ISBN 978-0-471-38461-8.
↑ Gauthier J. A. (1994): The diversification of the amniotes. In: D. R. Prothero and R. M. Schoch (ed.) Major Features of Vertebrate Evolution: 129 – 159. Knoxville, Tennessee: The Paleontological Society.
↑ аб The Reptile Database. // Посетен на 19 август 2010.
↑ reptile (animal): Behaviour. // Britannica.com. Посетен на 16 март 2010.
↑ Reptile and Amphibian Defense Systems. // Teachervision.fen.com. Посетен на 16 март 2010.
↑ Animal Planet: Ferocious Crocs. // Animal.discovery.com, 10 септември 2008. Посетен на 16 март 2010.
Литература |
- Laurin M & Gauthier J A. 1999. Phylogeny and classification of amniotes, Amniota (Tree of Life web project).
- Gauthier J A, Kluge A G, & Rowe T. 1988 г. The early evolution of the Amniota. In: Benton M J (ed.) The Phylogeny and Classification of the Tetrapods, Vol. 1: Amphibians, Reptiles, Birds, pp. 103 – 155. Оксфорд: Clarendon Press.
- Gauthier J A. 1994 г. The diversification of the amniotes. In: Prothero D R & Schoch R M (eds.) Major Features of Vertebrate Evolution, pp. 129 – 159. Ноксвил, Тенеси: The Paleontological Society.
- Германов А. Приложна зоология. Земиздат, 1992, ISBN 954-05-0187-3
- Пешев, Даниело. Зоология на гръбначните животни. Булвест 2000, 2002. ISBN 954-18-0278-8.
Вижте също |
- Херпетология
- Класификация на влечугите
- Влечугите в България
- Херпетофобия
Външни препратки |
- EMBL Reptile Database
Категория:
- Влечуги
(window.RLQ=window.RLQ||[]).push(function()mw.config.set("wgPageParseReport":"limitreport":"cputime":"1.184","walltime":"1.370","ppvisitednodes":"value":18017,"limit":1000000,"ppgeneratednodes":"value":35837,"limit":1500000,"postexpandincludesize":"value":148414,"limit":2097152,"templateargumentsize":"value":90926,"limit":2097152,"expansiondepth":"value":20,"limit":40,"expensivefunctioncount":"value":4,"limit":500,"unstrip-depth":"value":0,"limit":20,"unstrip-size":"value":19718,"limit":5000000,"entityaccesscount":"value":2,"limit":400,"timingprofile":["100.00% 1132.020 1 -total"," 70.07% 793.178 1 Шаблон:Taxobox"," 66.90% 757.351 1 Шаблон:Infobox"," 34.00% 384.905 1 Шаблон:Fossil_range"," 28.78% 325.841 1 Шаблон:Phanerozoic"," 26.52% 300.225 11 Шаблон:Fossil_range/bar"," 22.13% 250.467 34 Шаблон:Period_start"," 21.02% 237.904 14 Шаблон:Clade"," 16.29% 184.453 18 Шаблон:Cite"," 11.28% 127.711 1 Шаблон:Taxobox/authority"],"scribunto":"limitreport-timeusage":"value":"0.191","limit":"10.000","limitreport-memusage":"value":3274088,"limit":52428800,"cachereport":"origin":"mw1256","timestamp":"20190416050748","ttl":2592000,"transientcontent":false););"@context":"https://schema.org","@type":"Article","name":"u0412u043bu0435u0447u0443u0433u0438","url":"https://bg.wikipedia.org/wiki/%D0%92%D0%BB%D0%B5%D1%87%D1%83%D0%B3%D0%B8","sameAs":"http://www.wikidata.org/entity/Q10811","mainEntity":"http://www.wikidata.org/entity/Q10811","author":"@type":"Organization","name":"Contributors to Wikimedia projects","publisher":"@type":"Organization","name":"u0424u043eu043du0434u0430u0446u0438u044f u0423u0438u043au0438u043cu0435u0434u0438u044f","logo":"@type":"ImageObject","url":"https://www.wikimedia.org/static/images/wmf-hor-googpub.png","datePublished":"2005-01-23T23:58:36Z","dateModified":"2019-04-15T15:34:43Z","image":"https://upload.wikimedia.org/wikipedia/commons/5/5c/Total_internal_reflection_of_Chelonia_mydas.jpg","headline":"u043au043bu0430u0441 u0445u043eu0440u0434u043eu0432u0438 u0436u0438u0432u043eu0442u043du0438"(window.RLQ=window.RLQ||[]).push(function()mw.config.set("wgBackendResponseTime":155,"wgHostname":"mw1253"););


